
explain figure 2 please on the third page
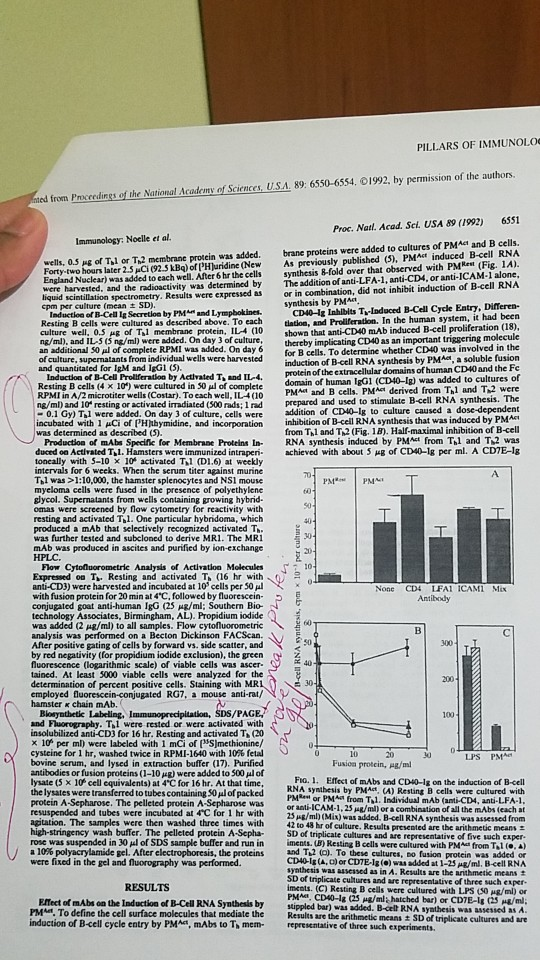
PILLARS OF IMMUNOLO ed from Proccedings of the National Academy of Sciences, U.S.A. 89: 6550-6554. 01992, by permission of the authors. 6551 Proc. Natl. Acad. Sel. USA 89 (1992) PMAct İnduced B cell RNA wells, 0.5 Ag of T,l or T,2 membrane protein was added. brane proteins were added to cultures of PMAct and B cells As previously published (5) Forty-tw hours later 2.5pC(92.5 kBq) ofPHJuridine(New England Nuclear) was added to each well. After 6 hr the cells and the radioactivity was determined by spectrometry. Results were expressed as synthesis &-fold over that observed with PMRet (Fig. 1A). The addition of anti-LFA-1, anti-CD4, or anti-ICAM-1 alone, or in combination, did not inhibit induction of B-cell RNA by Induction of B-Cell Ig Secretion by PMA and Lympbokines. Resting B cells were cultured as described above. To each CD40-Iz Inhibits T-Induced B-Cell Cycle Entry, Differen- latlon, and Proliferation. In the human system, it had been of Tl membrane protein, IL (10 shown that anti-CD40 mAb induced B-cell proliferation (18) culture well. ng/m), and IL ( ne/ml) were added. On day 3 of culture, thereby implicating CD40 as an important tnggering an additional 50 pl of complete RPMI was added. On day 6 for B cells. To determine whether CD40 was involved in the of culture, supernatants from individual wells were harvested Induction of B-Cell Proliferation by Activated Tand IL-4 Resting B cells (4 x 10") were cultured in 50 of complete RPMI in A/2 microtiter wells (Costar). To each well, IL4(10 protein of the extracellular domains of human CD40 and the Fe domain of human IgGI (CD40-1g) was added to cultures of PMA and B cells. PMAt derived from hl and T2 were ng/ml) and 10 resting or activated irradiated (500 rads; I rad prepared and used to stimulate B-cell RNA sy 0.1 Gy) Til were added. On day 3 of culture, cells were addition of CD40-1g to culture caused a inhibition of B-cell RNA synthesis that was induced by PMAt from Thl and Th2 (Fig. 18). Half-maximal inhibition of B-cell Production of mAbs Specific for Membrane Proteins In- RNA synthesis induced by PMct from Thl and T2 was duced oa Activated TI. Hamsters were immunized intraperi. achieved with about 5 ㎍ of CD4-Ig per ml. A CDIE-lg incubated with 1 Ci of PHjthymidine, and incorporation toncally with 5-10 x 10 activated Til (D1.6) at weekly intervals for 6 weeks. When the serum titer against murine Tl was >1:10,000, the hamster splenocytes and NS1 mouse myeloma cells were fused in the presence of polyethylene glycol. Supernatants from wells containing growing hybrid- resting and activated Tl. One particular hybridoma, which produced a mAb that selectively recognized activated T was further tested and subcloned to derive MR1. The MR mAb was produced in ascites and purified by ion-exchange of 10 Expressed o T Resting and activated T (16 hr with anti-CD3) were harvested and incubated at 10 cells per 50 l with fusion protein for 20 min at 4'C, followed by fluorescein conjugated goat anti-human IgG (25 ㎍/ml; Southern Bio- technology Associates, Birmingham, AL). Propidium iodide was added (2 μ8/ml) to all samples. Flow cytonuoro metric analysis was performed on a Becton Dickinson FACScan After positive gating of cells by forward vs. side scatter, and None CD4 IFAI ICAMI Mix by red negativity (for propidium iodide exclusion), the green o 2 fluorescence (logarithmic scale) of viable cells was ascer- tained. At least 5000 viable cells were analyzed for the determination of percent positive cells. Staining with MRL 30 employed fluorescein-conjugated RG7, a mouse anti-rat/ 100 and Fluorography. Til were rested or were activated with insolubilized anti-CD3 for 16 hr. Resting and activated Ti (20 x 10 per ml) were labeled with 1 mCi of (NS)methionine/ cysteine for l hr, washed twice in RPMI-1640 with 10% fetal bovine serum, and lysed in extraction buffer (17). Purified antibodies or fusion proteins (1-10 ㎍) were added to 500㎕ of lysate (5 x 10 cell equivalents) at 4'C for 16 hr. At that time, the lysates were transferred to tubes containing S0ul of packed protein A-Sepharose. The pelleted protein A-Sepharose was resuspended and tubes were incubated at 4"C for 1 hr with 10 Jo Fio. 1. Effect of mAbs and CD40-Ig on the induction of B-cell RNA synthesis by PMAct, (A) Resting B cells were cultured with PMRes or PM from T,l. Individual mAb (anti-CD4, anti-LFA-1 or anti-ICAM-1,25 ug/mi) or a combination of all the mAbs (each at 25Ag/ml) (Mix) was added. B-cell RNA synthesis was assessed from Results presented are the arithmetic means - of triplicate cultures and are representative of five such exper. on. The samples were then washed three times with 42 to 48 hr of culture. wash buffer. The pelleted protein A-Sepha S rose was suspended in 30 μ1 of SDS sample buffer and run in 10% polyacrylamide gel. After electropboresis, the proteins imens, un Resting B cells were cultured with PMA" fronta andT2 a. To these cultures, to fusion protein was added or were fixed in the gel and fluorography was performed CD40Ig(A, a) or CDTE-la (o) was added at 1-25s/ml. B-cell RNA synthesis was assessed as in A. Results are the arithmetic means t iments. (C) Resting B cells were cultured with LPS (S0 ug/ml) or PMAd. CD40-1g (25 ㎍/m, hatched bar) or CD7E-lg (25 ㎍/m2 Effect of mAbs on the Induction of B-Cell RNA Synthesis by stppled bar) wa PMA. To define the cell surface molecules that mediate the Results are the induction of B-cell cycle entry by PMA, mAbs to Th mem- epresentative of three such experiments stippled bar) was added. B-cell RNA syathesis was assessed as A arithmetic means SD of triplicate cultures and are
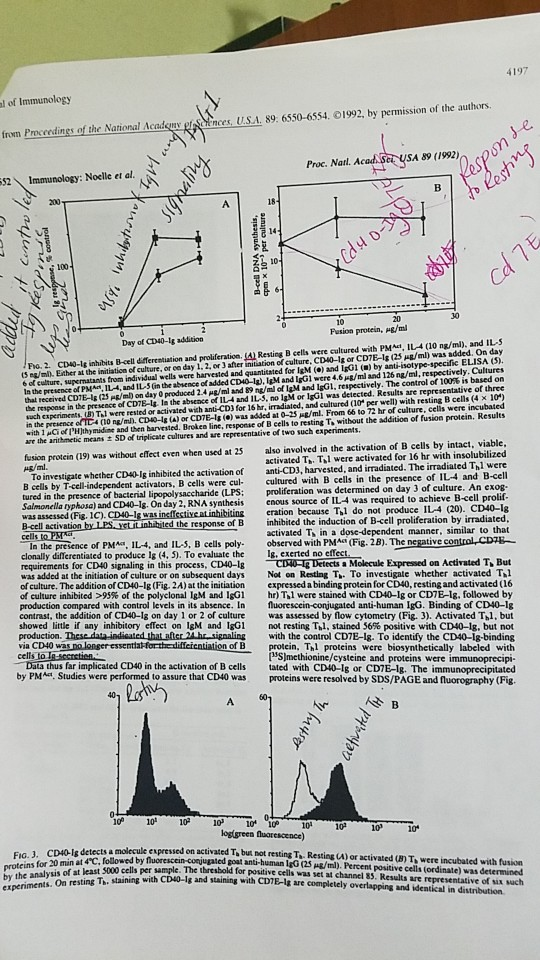
fl ner.U.SA 89 6550-6554. С 1992, by permission of the authors ader fron Pro erdings of the National Proc. Natl. Acad Ser USA 89 (1992) 52Immunology: Noelle et al. and ILS S e/mil Ether at the initiation ef culture, or eo day 1. 2, or 3 after initiation of culture. CD40-1g or CDTE-Is (23 Hg/ml) was added. On day 6 of culture, supernatants trom individual wells were harvested and quanticated for IsM () and IgG1 () by anti-isotype-specific ELISA 3) In the presence of PMA, 114, and IL-5 Gn the absence of added CDO-la. IgM and IgG1 were 4.6 ㎍/ml and i 26 ng/m, respectively. Cultures thal received CD18-lg (25 Pg/ml) on day 0 produced 2.4 ㎍/ml and 89 ng/ml of IgM and igG1, respectively. The control of 100% is based on the response in the presence of CD7E-ls la the abseace of IL4 and IL-5, no lgM or IG1 was detected, Results are representative of three chexperiments un Tal were rested or activated with ant-CD) for ishr, imdialed, and cultured (10, per well) with resting B cells (4 10) in the presence of TC4 (10 ng mn CD#4g (A) or CD7E-1 .) was added at 0-25 ㎍/mL From 66 to 72 hr of culture, cells were incubated with 1 Cof lH)thy midime and then harvested. Brokeo line, response of B cells to resting To without the addition of fusion protein. Results / Po. 2. cD0-Is inhabits B-ell fferestiuation and proliíteration ()Resting B cells were cultured with PM, arr the arithmetic means ± SD of triplicate altures and are representative er two such apenmenis. protein (19) was without effect even when used at 25s also involved in the activation of B cells by intact, viabl B cells by T-cell-independent activators, B cells were cul cultured with B Salmonella typhosa) and CD40-Is. On day 2,. RNA synthesis enous source of activated TTl were activated for 16 hr with insolubilized To investigate whether CD40-lg inhibited the activation of anti-CD3, harvested, and irradiated. The irradiated Thl were cells in the presence of IL4 and B-cell on was determined on day 3 of culture. An exog was assessed (Fis. 1C). CD40-g eration because Thl do not produce IL4 (20). CD40-lg response of B inhibited the induction of B-cell proliferation by irradiated, activated T in a dose-dependent manner, similar to that of PMA IL-4, and IL, B cells poly. observed with PMA (Fig. 2B). The negative control CDTE- Ig, exerted no eff clonally differentiated to produce Ig (4, 5). To evaluate the requirements for CD40 signaling in this process, CD40-1s was added at the initiation of culture or on subsequent days of culture. The addition of CD40-1g (Fig. 2A) at the initiation expressed a binding protein for CD40, resting and activated (16 of culture inhibited >99% of the polyclonal IgM and IgG1 hr) Ta were stained with CD40-lg or CDIE-lg, followed by production compared with control levels in its absence. In fluorescein-conjugated anti-human IgG. Binding of CD40-1g contrast, the addition of CD40-1g on day 1 or 2 of culture was assessed by flow cytometry (Fig. 3). Activated Thl, but showed little if any inhibitory effect on IgM and IgG1 production. These dataindisated that after via CD40 was no lonser essentiakforth a Molecule Expressed on Activated T, But Not on Resting T. To investigate whether activated T.l not resting Tal, stained 56% positive with CD40-Ig, but not 24he signaling with the control CD7E-lg. To identify the CD40-1g-binding protein, Thl proteins were biosynthetically labeled with of B ed CD4Oin the activation of B cells by PMa, Studies were performed to assure that CD40 was SJmethionine/cysteine and proteins were tated with CDW-Ig or CDE-lg·The immunoprecipitated proteins were resolved by Sps/PAGE and fuorography (Fig. 10 10 10 10 10 010 10 logigreen proteins for 20 min at 4°C, followed by by the analysis of at least 5000 cells per sample. experiments. On resting Th, staining on activated Tabu·thot resting Ta-Resting U) or activated un Th were incubaled with thia ample. The thresbold for positive cells was set at channel 35. Results are representative of six sach with CD40-1g and staining with CDTE-g are completely overlapping and identical in distribution
Homework Answers
In figure 2.A - shown that CD inhibit T-cell independent (without T cell), B cell activation. when B cell are culture with (PMact,IL-4 and IL-5) CD40-Ig at initial period, their is approximately 95 % inhibition is occur in IgG1 and IgM synthesis or poly clonal Ab production , but if CD40-Ig applied in 2 or 3 day of B cell culture then their is very less or no inhibitor effect is seen in both antibody production. While in control solution were CD40-Ig is absent their is normal production of poly clonal Ab from B cell through independent manner, due to the presence of PMactand (IL-4 and IL-5).
In figure 2B.Indicate CD-40 help B cell to active from fully intact T cell , IL-4 and IL-5 not from the radiation influence T cell. In control conditions were CD-40 Ig is absent and other are same conditions but their is decrease B cell proliferation is seen (negative control) because of distract T cell interaction.
So the Role of CD-40 Ig is to insure B cell activate through intact T cell only (T dependent activation only).
Add Answer to:
The Jounal of Immunol nes of the National Academy of Sciences USA 89. 6550-6554. 01992, by permis...
immunology practing samples and not sure if my answers correct. 1. NK cells are effective against...
 immunology
practing samples and not sure if my answers correct.
1. NK cells are effective against viral infections because many virus infected cells a. are susceptible to lysis b. are susceptible to phagocytosis c. show reduced levels of MHC class I molecules d. show reduced levels of MHC class II molecules e. none of the above 2. Arachidonic acid serves as a substrate in the lipoxygenase and cyclooxygenase pathways resulting in the production of a. histamine, leukotrienes b. leukotrienes, prostaglandins...
immunology
practing samples and not sure if my answers correct.
1. NK cells are effective against viral infections because many virus infected cells a. are susceptible to lysis b. are susceptible to phagocytosis c. show reduced levels of MHC class I molecules d. show reduced levels of MHC class II molecules e. none of the above 2. Arachidonic acid serves as a substrate in the lipoxygenase and cyclooxygenase pathways resulting in the production of a. histamine, leukotrienes b. leukotrienes, prostaglandins...
Q 4. Explain how Fig.1A (arrow) result corroborates with the author hypothesis about Parkin aggre...
 Q
4. Explain how Fig.1A (arrow) result corroborates with the author
hypothesis about Parkin aggresome and proteasome activity in PD?
MG-132+ HMW 127- 84-- 41- DTT+ IP T HMW 6- IP : anti-flag WB: anti-ubiquitin Fia. 1. Parkin forms plexes A, HEK 293Tells transiently tranafected with FLAG-Parkin were divided into twe dishes and were either untreated or treated with 5PM MG-132 for 161). Cells were then lysed abidใหr-taining 1% Triton X-100 ลnd fractitated into "Iuble (S) and insoluble u) frac...
Q
4. Explain how Fig.1A (arrow) result corroborates with the author
hypothesis about Parkin aggresome and proteasome activity in PD?
MG-132+ HMW 127- 84-- 41- DTT+ IP T HMW 6- IP : anti-flag WB: anti-ubiquitin Fia. 1. Parkin forms plexes A, HEK 293Tells transiently tranafected with FLAG-Parkin were divided into twe dishes and were either untreated or treated with 5PM MG-132 for 161). Cells were then lysed abidใหr-taining 1% Triton X-100 ลnd fractitated into "Iuble (S) and insoluble u) frac...
Please summarise the following content:(no word limit) EXPERIMENTAL PROCEDURES Animals All animals including pregnant ICR mice,...
 Please summarise the following content:(no word
limit)
EXPERIMENTAL PROCEDURES Animals All animals including pregnant ICR mice, neonatal Wistar rats, and NOD-SCID mice (8 weeks old, male) were purchased from CLEA Japan (Tokyo). All exper- imental procedures and protocols were approved by the Animal Care and Use Committee of Keio University and conformed to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Cells Mouse ESCs were obtained from the Laboratory of Pluripotent Cell Studies, RIKEN...
Please summarise the following content:(no word
limit)
EXPERIMENTAL PROCEDURES Animals All animals including pregnant ICR mice, neonatal Wistar rats, and NOD-SCID mice (8 weeks old, male) were purchased from CLEA Japan (Tokyo). All exper- imental procedures and protocols were approved by the Animal Care and Use Committee of Keio University and conformed to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Cells Mouse ESCs were obtained from the Laboratory of Pluripotent Cell Studies, RIKEN...
BELOW IS A METHOD. WHAT PROTOCOL (ALSO BELOW: A-F) MATCHES THIS METHOD?
 BELOW IS A METHOD. WHAT PROTOCOL (ALSO BELOW: A-F)
MATCHES THIS METHOD?
Method 5: Controlled ovarian stimulation and oocyte recovery was performed. Briefly, cycling females were subjected to follicular stimulation using twice-daily intramuscular injections of recombinant human FSH as well as concurrent treatment with Antide, a GnRH antagonist, for days 8-9. Unless indicated otherwise, all reagents were from Sigma-Aldrich and all hormones were from Ares Advanced Technologies Inc. Females received recombinant human luteinizing hormone on days 7-9 and recombinant...
BELOW IS A METHOD. WHAT PROTOCOL (ALSO BELOW: A-F)
MATCHES THIS METHOD?
Method 5: Controlled ovarian stimulation and oocyte recovery was performed. Briefly, cycling females were subjected to follicular stimulation using twice-daily intramuscular injections of recombinant human FSH as well as concurrent treatment with Antide, a GnRH antagonist, for days 8-9. Unless indicated otherwise, all reagents were from Sigma-Aldrich and all hormones were from Ares Advanced Technologies Inc. Females received recombinant human luteinizing hormone on days 7-9 and recombinant...
1. The prevention of inflammatory immune responses to inhaled antigens in healthy individuals has mechanisms in...
1. The prevention of inflammatory immune responses to inhaled antigens in healthy individuals has mechanisms in common with those that prevent inflammatory immune responses to commensal microbes in the gut. One important component of immune regulation shared by these two situations is: a. The induction of increased numbers of IFN-g-producing T cells and ILCs in the airway and gastrointestinal epithelium b. The presence of tissue-resident mast cells that bind IgE through the high affinity IgE receptor c. The high levels...
For background: Q6= tyrosine kinase inhibitors & Q8= MHC molecules I need help with Q7 &...
 For background: Q6= tyrosine kinase inhibitors & Q8= MHC
molecules
I need help with Q7 & Q9. I think Q7 is C but not sure. For
Q9 I think it is phorbol myristate acetate since in my notes it
says "Treatment of PMA and ionomycin on T cells can induce the
effects of TCR stimulation" does this reasoning make sense?
Although the (Q-6) approaches of CML treatment have dramatically changed the treatment of CML by inducing long term overall survival...
For background: Q6= tyrosine kinase inhibitors & Q8= MHC
molecules
I need help with Q7 & Q9. I think Q7 is C but not sure. For
Q9 I think it is phorbol myristate acetate since in my notes it
says "Treatment of PMA and ionomycin on T cells can induce the
effects of TCR stimulation" does this reasoning make sense?
Although the (Q-6) approaches of CML treatment have dramatically changed the treatment of CML by inducing long term overall survival...
PLEASE COMPUTER TYPED* OR VERY CLEAR HANDWRITING with details 1. Radioactive (14C) uridine (lots of it)...
 PLEASE COMPUTER TYPED* OR VERY CLEAR HANDWRITING with
details
1. Radioactive (14C) uridine (lots of it) is added to a growing culture of bacteria for 10 seconds and then the drug rifamycin is added (rifamycin blocks the initiation of transcription, but not continued elongation of RNA polymerases that have already begun transcribing). At two-minute intervals, 1 ml samples are removed and placed on ice. The bacteria are lysed (broken open) and an enzyme is added that will chew up all...
PLEASE COMPUTER TYPED* OR VERY CLEAR HANDWRITING with
details
1. Radioactive (14C) uridine (lots of it) is added to a growing culture of bacteria for 10 seconds and then the drug rifamycin is added (rifamycin blocks the initiation of transcription, but not continued elongation of RNA polymerases that have already begun transcribing). At two-minute intervals, 1 ml samples are removed and placed on ice. The bacteria are lysed (broken open) and an enzyme is added that will chew up all...
1. You are studying phage transduction and trying to deliver a kanamycin resistance gene from one...
 1. You are studying phage transduction and trying to deliver a kanamycin resistance gene from one strain of bacteria to another using phage. The kanR gene is FAR away from the lambda prophage in your bacterium. Choose which type of transduction should you use for your experiment and explain why. a. lambda phage, specialized transduction b. lambda phage, conjugation C. T4 phage, generalized transduction d. A different phage with mitomycin C induction 2. A pig is infected with two subtypes...
1. You are studying phage transduction and trying to deliver a kanamycin resistance gene from one strain of bacteria to another using phage. The kanR gene is FAR away from the lambda prophage in your bacterium. Choose which type of transduction should you use for your experiment and explain why. a. lambda phage, specialized transduction b. lambda phage, conjugation C. T4 phage, generalized transduction d. A different phage with mitomycin C induction 2. A pig is infected with two subtypes...
lab question 1. What is the basis of the different purification methods? 2. What are some...
lab question 1. What is the basis of the different purification methods? 2. What are some of the factors the might have interfered with your results? 3. How might you improve the process to increase the yield and purity? lab process E. coli BL21 (DE3) cells were transformed with the pET Topo-1521 vector containing a reading frame encoding the green fluorescent protein (GFP). Cells were cultured in M9ZB media at 37°C until the absorbance at 600 nm reached 0.7, at...
SHORT ANSWER. Write the word or phrase that best completes each statement or answers the question....
 SHORT ANSWER. Write the word or phrase that best completes each statement or answers the question. 00 OA Figure 2.1 Using Figure 2.1, match the following: 1) Tertiary (protein) structure, 2) Monosaccharide 3) Nucleotide 4) Lipid 5) Functional protein 6) Polymer 7) Polysaccharide TRUE FALSE. Write T if the statement is true and F if the statement is false, 8) Final preparation for cell division is made during the cell life cycle subphase called G2. SHORT ANSWER. Write the word...
SHORT ANSWER. Write the word or phrase that best completes each statement or answers the question. 00 OA Figure 2.1 Using Figure 2.1, match the following: 1) Tertiary (protein) structure, 2) Monosaccharide 3) Nucleotide 4) Lipid 5) Functional protein 6) Polymer 7) Polysaccharide TRUE FALSE. Write T if the statement is true and F if the statement is false, 8) Final preparation for cell division is made during the cell life cycle subphase called G2. SHORT ANSWER. Write the word...
 immunology
practing samples and not sure if my answers correct.
1. NK cells are effective against viral infections because many virus infected cells a. are susceptible to lysis b. are susceptible to phagocytosis c. show reduced levels of MHC class I molecules d. show reduced levels of MHC class II molecules e. none of the above 2. Arachidonic acid serves as a substrate in the lipoxygenase and cyclooxygenase pathways resulting in the production of a. histamine, leukotrienes b. leukotrienes, prostaglandins...
immunology
practing samples and not sure if my answers correct.
1. NK cells are effective against viral infections because many virus infected cells a. are susceptible to lysis b. are susceptible to phagocytosis c. show reduced levels of MHC class I molecules d. show reduced levels of MHC class II molecules e. none of the above 2. Arachidonic acid serves as a substrate in the lipoxygenase and cyclooxygenase pathways resulting in the production of a. histamine, leukotrienes b. leukotrienes, prostaglandins...
 Q
4. Explain how Fig.1A (arrow) result corroborates with the author
hypothesis about Parkin aggresome and proteasome activity in PD?
MG-132+ HMW 127- 84-- 41- DTT+ IP T HMW 6- IP : anti-flag WB: anti-ubiquitin Fia. 1. Parkin forms plexes A, HEK 293Tells transiently tranafected with FLAG-Parkin were divided into twe dishes and were either untreated or treated with 5PM MG-132 for 161). Cells were then lysed abidใหr-taining 1% Triton X-100 ลnd fractitated into "Iuble (S) and insoluble u) frac...
Q
4. Explain how Fig.1A (arrow) result corroborates with the author
hypothesis about Parkin aggresome and proteasome activity in PD?
MG-132+ HMW 127- 84-- 41- DTT+ IP T HMW 6- IP : anti-flag WB: anti-ubiquitin Fia. 1. Parkin forms plexes A, HEK 293Tells transiently tranafected with FLAG-Parkin were divided into twe dishes and were either untreated or treated with 5PM MG-132 for 161). Cells were then lysed abidใหr-taining 1% Triton X-100 ลnd fractitated into "Iuble (S) and insoluble u) frac...
 Please summarise the following content:(no word
limit)
EXPERIMENTAL PROCEDURES Animals All animals including pregnant ICR mice, neonatal Wistar rats, and NOD-SCID mice (8 weeks old, male) were purchased from CLEA Japan (Tokyo). All exper- imental procedures and protocols were approved by the Animal Care and Use Committee of Keio University and conformed to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Cells Mouse ESCs were obtained from the Laboratory of Pluripotent Cell Studies, RIKEN...
Please summarise the following content:(no word
limit)
EXPERIMENTAL PROCEDURES Animals All animals including pregnant ICR mice, neonatal Wistar rats, and NOD-SCID mice (8 weeks old, male) were purchased from CLEA Japan (Tokyo). All exper- imental procedures and protocols were approved by the Animal Care and Use Committee of Keio University and conformed to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Cells Mouse ESCs were obtained from the Laboratory of Pluripotent Cell Studies, RIKEN...
 BELOW IS A METHOD. WHAT PROTOCOL (ALSO BELOW: A-F)
MATCHES THIS METHOD?
Method 5: Controlled ovarian stimulation and oocyte recovery was performed. Briefly, cycling females were subjected to follicular stimulation using twice-daily intramuscular injections of recombinant human FSH as well as concurrent treatment with Antide, a GnRH antagonist, for days 8-9. Unless indicated otherwise, all reagents were from Sigma-Aldrich and all hormones were from Ares Advanced Technologies Inc. Females received recombinant human luteinizing hormone on days 7-9 and recombinant...
BELOW IS A METHOD. WHAT PROTOCOL (ALSO BELOW: A-F)
MATCHES THIS METHOD?
Method 5: Controlled ovarian stimulation and oocyte recovery was performed. Briefly, cycling females were subjected to follicular stimulation using twice-daily intramuscular injections of recombinant human FSH as well as concurrent treatment with Antide, a GnRH antagonist, for days 8-9. Unless indicated otherwise, all reagents were from Sigma-Aldrich and all hormones were from Ares Advanced Technologies Inc. Females received recombinant human luteinizing hormone on days 7-9 and recombinant...
 For background: Q6= tyrosine kinase inhibitors & Q8= MHC
molecules
I need help with Q7 & Q9. I think Q7 is C but not sure. For
Q9 I think it is phorbol myristate acetate since in my notes it
says "Treatment of PMA and ionomycin on T cells can induce the
effects of TCR stimulation" does this reasoning make sense?
Although the (Q-6) approaches of CML treatment have dramatically changed the treatment of CML by inducing long term overall survival...
For background: Q6= tyrosine kinase inhibitors & Q8= MHC
molecules
I need help with Q7 & Q9. I think Q7 is C but not sure. For
Q9 I think it is phorbol myristate acetate since in my notes it
says "Treatment of PMA and ionomycin on T cells can induce the
effects of TCR stimulation" does this reasoning make sense?
Although the (Q-6) approaches of CML treatment have dramatically changed the treatment of CML by inducing long term overall survival...
 PLEASE COMPUTER TYPED* OR VERY CLEAR HANDWRITING with
details
1. Radioactive (14C) uridine (lots of it) is added to a growing culture of bacteria for 10 seconds and then the drug rifamycin is added (rifamycin blocks the initiation of transcription, but not continued elongation of RNA polymerases that have already begun transcribing). At two-minute intervals, 1 ml samples are removed and placed on ice. The bacteria are lysed (broken open) and an enzyme is added that will chew up all...
PLEASE COMPUTER TYPED* OR VERY CLEAR HANDWRITING with
details
1. Radioactive (14C) uridine (lots of it) is added to a growing culture of bacteria for 10 seconds and then the drug rifamycin is added (rifamycin blocks the initiation of transcription, but not continued elongation of RNA polymerases that have already begun transcribing). At two-minute intervals, 1 ml samples are removed and placed on ice. The bacteria are lysed (broken open) and an enzyme is added that will chew up all...
 1. You are studying phage transduction and trying to deliver a kanamycin resistance gene from one strain of bacteria to another using phage. The kanR gene is FAR away from the lambda prophage in your bacterium. Choose which type of transduction should you use for your experiment and explain why. a. lambda phage, specialized transduction b. lambda phage, conjugation C. T4 phage, generalized transduction d. A different phage with mitomycin C induction 2. A pig is infected with two subtypes...
1. You are studying phage transduction and trying to deliver a kanamycin resistance gene from one strain of bacteria to another using phage. The kanR gene is FAR away from the lambda prophage in your bacterium. Choose which type of transduction should you use for your experiment and explain why. a. lambda phage, specialized transduction b. lambda phage, conjugation C. T4 phage, generalized transduction d. A different phage with mitomycin C induction 2. A pig is infected with two subtypes...
 SHORT ANSWER. Write the word or phrase that best completes each statement or answers the question. 00 OA Figure 2.1 Using Figure 2.1, match the following: 1) Tertiary (protein) structure, 2) Monosaccharide 3) Nucleotide 4) Lipid 5) Functional protein 6) Polymer 7) Polysaccharide TRUE FALSE. Write T if the statement is true and F if the statement is false, 8) Final preparation for cell division is made during the cell life cycle subphase called G2. SHORT ANSWER. Write the word...
SHORT ANSWER. Write the word or phrase that best completes each statement or answers the question. 00 OA Figure 2.1 Using Figure 2.1, match the following: 1) Tertiary (protein) structure, 2) Monosaccharide 3) Nucleotide 4) Lipid 5) Functional protein 6) Polymer 7) Polysaccharide TRUE FALSE. Write T if the statement is true and F if the statement is false, 8) Final preparation for cell division is made during the cell life cycle subphase called G2. SHORT ANSWER. Write the word...
Most questions answered within 3 hours.
-
As part of the process for improving the quality of their cars,
Toyota engineers have identified...
asked 8 minutes ago -
1. You have genotyped an entire population of lab mice for a
single locus (let's label...
asked 5 minutes ago -
What is the relationship between the number of coils in a
solenoid and the emf induced...
asked 33 minutes ago -
cell in human body reproduces. what process does it use? describe
the steps in the cell...
asked 1 hour ago -
The cheetah is one of the fastest accelerating animals, for it
can go from rest to...
asked 2 hours ago -
CaCO3 (s) ⇌ CaO (s) + CO2 (g)
At 637 °C, the reaction reaches equilibrium.
If...
asked 3 hours ago -
The condensate from a steam distillation contains 12.0 g of
compound A and 18.0 g of...
asked 6 hours ago -
A major economic benefit of fixed exchange rates compared to
floating rates is that
a. the...
asked 6 hours ago -
1) For this reaction,
SiCl4 (l)+ 2H2O(g) <->
SiO2 (s) + 4HCl (g) DeltaH = -127 KJ...
asked 6 hours ago -
Which of the following does NOT add to US GDP? A. Saudi Arabia
buys fighter jets...
asked 8 hours ago -
2. Describe market equilibrium in terms of the following
characteristics
d.
How supply and demand interactions...
asked 7 hours ago -
1a. Create a class named Computer
- Separate declaration from implementation (i.e. Header and CPP
files)...
asked 8 hours ago