Using one example of each class of gene, explain how mutations in tumour suppressor genes and...
Using one example of each class of gene, explain how mutations in tumour suppressor genes and oncogenes can lead to alterations in cell signalling pathways, and contribute to cancer development.
Homework Answers
The mutations incurred in the normal-functioning genes can be classified as
- Gain of function mutation: mutation leading to the ability of new functions suppressing the normal functioning of the gene.
- Loss of function mutation: mutation leading to losing the ability of the gene to perform the normal functioning.
Mutations in TP53 - Tumor suppressor gene
TP53 encodes the p53 protein, an important tumor suppressor protein. Many different types of cancer show a high incidence of TP53 mutations, leading to the expression of mutant p53 proteins. TP53 is the most commonly mutated gene in human cancer. Alterations have been found in virtually every region of the protein, but only a handful of the most frequently occurring mutations have been studied in depth for their contribution to cancer progression. In some cases, frameshift or nonsense mutations result in the loss of p53 protein expression, as seen with other tumor suppressors. However, more frequently, the tumor-associated alterations in p53 result in missense mutations, leading to the substitution of a single amino acid in the p53 protein that can be stably expressed in the tumor cell. These substitutions occur throughout the p53 protein, but most commonly cluster within the DNA binding region of p53, with six “hotspot” amino acids that are most frequently substituted. These mutations generally lead to a loss or diminution of the wild-type activity of p53, and because p53 normally acts as a tetramer, these mutant proteins may also function as dominant negative inhibitors over any remaining wild-type p53.
p53 is a transcription factor able to regulate several intracellular pathways involved in cell survival, DNA-repair, apoptosis, and senescence. p53 is also known as “the guardian of the genome”, being able to preserve DNA integrity in response to a number of stimuli, such as ionizing radiations, genotoxic insults and oxidative stress. Several studies have demonstrated that many mutant p53 proteins, not only lose their tumor suppression functions but also gain new oncogenic functions. This phenomenon is termed “the gain of function of mutant p53”. More specifically, mutant p53 interacts with proteins that normally partner with wild type p53. This new association deprives them of their anticancer activities and in place, they are corrupted to act as cancerogenesis promoters.
PML stabilizes and activates wild type p53 only in presence of stressed conditions, while, when p53 is mutated its association with PML is constitutive. Mutant p53-PML stabilized interaction leads p53 to aberrantly transcribe its targets. The major consequences of the gain of functions mutant p53 affect the metabolism and the response to oxidative stress of cancer cells. Rapid cell proliferation strongly needs a ready supply of energy and basic macromolecules, so mutant p53 proteins have been reported to facilitate the supply of cancer cells.
Mutant p53 is able to modify lipid metabolism in a way that encourages rapid cell proliferation and intracellular signaling. In fact, mutant p53 engages the SREBPs (sterol regulatory element-binding protein) and leads to FAs synthesis, β-oxidation arrest, and cholesterol synthesis, favoring so, the generation of a robust supply of lipids for cell membranes formation.
The strongest effect of mutant p53 on the metabolism regards the glucose. Mutant p53 constitutively stimulates glycolysis acting in various ways. First of all, it up-regulates GLUT1 and GLUT 4 favoring the rapid uptake of glucose in tumor cells, moreover, mutant p53 stimulated genes involved in glycolysis activation such as TIGAR, and finally, it stimulates the PPP acting on G6PD. The final result is a wide supply of glucose which can be consumed in order to produce energy also in aerobic conditions.
Rapid cell proliferation produces high levels of ROS, which may dampen cell membranes, protein structures and DNA inducing apoptosis. Mutant p53, as previously seen, can constitutively stimulate genes involved in antioxidant cell response. The greater response to the activation of these genes is the synthesis of molecules able to neutralize ROS, such as glutathione.


Mutations in Ras - proto-oncogene
Proto-oncogenes are a group of genes that cause normal cells to become cancerous when they are mutated. Mutations in proto-oncogenes are typically dominant in nature, and the mutated version of a proto-oncogene is called an oncogene. Often, proto-oncogenes encode proteins that function to stimulate cell division, inhibit cell differentiation, and halt cell death. All of these processes are important for normal human development and for the maintenance of tissues and organs. Oncogenes, however, typically exhibit increased production of these proteins, thus leading to increased cell division, decreased cell differentiation, and inhibition of cell death; taken together, these phenotypes define cancer cells.
All mammalian cells express three closely related Ras proteins: H-Ras, K-Ras, and N-Ras that promote oncogenesis when mutationally activated at codons 12, 13 or 61. Despite a high degree of similarity between the isoforms, K-Ras mutations are far more frequently observed in cancer and each isoform displays preferential coupling to particular cancer types. Ras proteins are proto-oncogenes that are frequently mutated in human cancers. They are encoded by three ubiquitously expressed genes: HRAS, KRAS, and NRAS. These proteins are GTPases that function as molecular switches regulating pathways responsible for proliferation and cell survival. Ras proteins are normally tightly regulated by guanine nucleotide exchange factors (GEFs) promoting GDP dissociation and GTP binding and GTPase-activating proteins (GAPs) that stimulate the intrinsic GTPase activity of Ras to switch off the signaling. Aberrant Ras function is associated with hyper-proliferative developmental disorders and cancer and in tumors is associated with a single mutation typically at codons 12, 13 or 61.
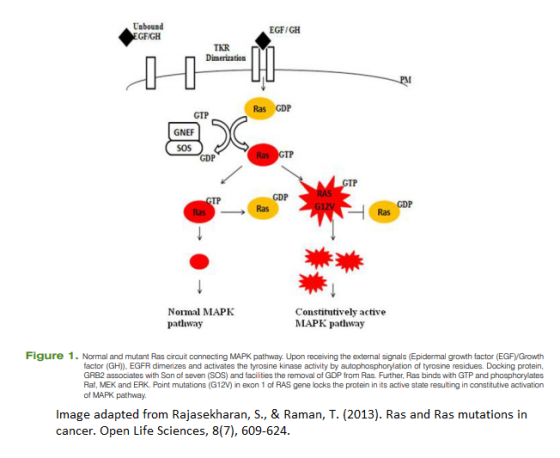
One of the most frequent ways in which the MAPK is set to overdrive is by a mutation in the RAS protein; mutations in the KRAS protein are found in about 40% of all colorectal cancer (CRC) cases. These mutations are sometimes present in early adenomas and in cells with the minimal potential to develop a malignancy. In molecular terms, mutations in these three KRAS/NRAS codons may lead to conformational changes so that the RAS-GAP protein cannot activate the inherent GTPase enzyme anymore. As a result, the GTP molecules are not hydrolyzed and instead they maintain RAS continuously in its active state, thus causing protumorigenic effects by amplifying signaling in the MAPK pathway.
Add Answer to:
Using one example of each class of gene, explain how mutations
in tumour suppressor genes and...
Cancer and Gene Regulation Why is a cell cycle control system needed for cell division? What...
 Cancer and Gene Regulation Why is a cell cycle control system needed for cell division? What happens when cells do NOT respond to the cell cycle control system and divide excessively? Tumor Proto-oncogeno (for protein that stimulates coll division) 6 Y DNA Benign Tumor= Mutation withln a control region of DNA Malignant Tumor Mutated promoter Metastasis Normal growth-stimulating protein in excess Oncogene Tumor-Suppressor Genes Proto-oncogene utled tara gese Samor-auppresr gane Many proto-oncogenes code for growth factors /Deletive nonimenig Normel grewt...
Cancer and Gene Regulation Why is a cell cycle control system needed for cell division? What happens when cells do NOT respond to the cell cycle control system and divide excessively? Tumor Proto-oncogeno (for protein that stimulates coll division) 6 Y DNA Benign Tumor= Mutation withln a control region of DNA Malignant Tumor Mutated promoter Metastasis Normal growth-stimulating protein in excess Oncogene Tumor-Suppressor Genes Proto-oncogene utled tara gese Samor-auppresr gane Many proto-oncogenes code for growth factors /Deletive nonimenig Normel grewt...
Which of the following is a false statements about tumor suppressor genes? Gene amplification (duplication) of...
Which of the following is a false statements about tumor suppressor genes? Gene amplification (duplication) of a tumor suppressor gene is less likely to result in cancer than gene amplification of a proto-onocogene. Individuals with a single normal copy of a tumor suppressor gene are more prone to cancer than those with two normal copies. Inactivation of tumor suppressor genes can lead to enhanced cell survival and cell proliferation. Epigenetic changes that silence tumor suppressor genes would not lead to...
BRCA1 and BRCA2 are tumor suppressor genes that have been associated with breast cancer development due...
BRCA1 and BRCA2 are tumor suppressor genes that have been associated with breast cancer development due to mutations of these genes being linked to a subset of breast cancers. If an individual carries one nonfunctional copy of the BRCA gene, will all of their cells be affected? Explain.
please help me with this genetics question! thank you! TS =Tumor Suppressor genes O = Oncogenes...
 please help me with this genetics question! thank you!
TS =Tumor Suppressor genes
O = Oncogenes
5A. (12pts) Cancer is a genetic disease. Some of the causative mutations are Tumor Suppressor genes, and others convert proto-oncogenes into Oncogenes. In the list of properties below, mark an X in the column for TS, O, or both. TS O both Causes of inherited elevated cancer risk. Leads to uncontrolled cell growth. Typically, spontaneous, gain-of-function mutations. Can cause increased DNA damage. Dominant in...
please help me with this genetics question! thank you!
TS =Tumor Suppressor genes
O = Oncogenes
5A. (12pts) Cancer is a genetic disease. Some of the causative mutations are Tumor Suppressor genes, and others convert proto-oncogenes into Oncogenes. In the list of properties below, mark an X in the column for TS, O, or both. TS O both Causes of inherited elevated cancer risk. Leads to uncontrolled cell growth. Typically, spontaneous, gain-of-function mutations. Can cause increased DNA damage. Dominant in...
Consider a spontaneous mutation in the gene HER2, a tumor suppressor involved in cell cycle control...
 Consider a spontaneous mutation in the gene HER2, a tumor suppressor involved in cell cycle control Mutations happen at a rate of about 1 error per 109 bases per generation. A spontaneous deleterious mutation must occur in HER2 specifically, and not in another gene. The likelihood that a spontaneous mutation is deleterious and could lead to cancer depends on the number of mutable bases in HER2, the fraction of bases that will affect the function of HER2, and that both...
Consider a spontaneous mutation in the gene HER2, a tumor suppressor involved in cell cycle control Mutations happen at a rate of about 1 error per 109 bases per generation. A spontaneous deleterious mutation must occur in HER2 specifically, and not in another gene. The likelihood that a spontaneous mutation is deleterious and could lead to cancer depends on the number of mutable bases in HER2, the fraction of bases that will affect the function of HER2, and that both...
Please help with answers to these questions answer the ones that are straightforward like this one...
 Please help with answers to these questions
answer the ones that are straightforward like this one
Question 24 5 pts 1. Beta-catenin is classified as a proto-oncogene and APC is classified as a tumor suppressor gene. Both are important parts of the Wnt signaling pathway. Describe the relationship between beta-catenin and APC. Explain in detail why they are classified as a proto-oncogene or tumor suppressor gene. 2. Suppose you're working in a cancer research lab and discover a small molecule...
Please help with answers to these questions
answer the ones that are straightforward like this one
Question 24 5 pts 1. Beta-catenin is classified as a proto-oncogene and APC is classified as a tumor suppressor gene. Both are important parts of the Wnt signaling pathway. Describe the relationship between beta-catenin and APC. Explain in detail why they are classified as a proto-oncogene or tumor suppressor gene. 2. Suppose you're working in a cancer research lab and discover a small molecule...
Discuss mechanisms by which gene expression may be altered. How do these alterations induce cancer-causing mutations...
Discuss mechanisms by which gene expression may be altered. How do these alterations induce cancer-causing mutations in cell DNA? Explain how cancer is formed.
BRCAll is a tumor suppressor gene located on chromosome pair 13. Inherited mutations in BRCAII can...
 BRCAll is a tumor suppressor gene located on chromosome pair 13. Inherited mutations in BRCAII can predispose an individual to breast cancer. If Scarlet carries an inherited mutation for BRCAII and her husband Scott does not, what is the probability that their son inherited the mutated allele? O 0% o 25% O 50% O 75% O 100% Question 2 1 pts A plasmid SELECT ALL THAT APPLY. is a small circular piece of DNA independent of the chromosome is a...
BRCAll is a tumor suppressor gene located on chromosome pair 13. Inherited mutations in BRCAII can predispose an individual to breast cancer. If Scarlet carries an inherited mutation for BRCAII and her husband Scott does not, what is the probability that their son inherited the mutated allele? O 0% o 25% O 50% O 75% O 100% Question 2 1 pts A plasmid SELECT ALL THAT APPLY. is a small circular piece of DNA independent of the chromosome is a...
Given that TP53 is a recessive gene and is not located on the X chromosome, why would people who inherit just one mutant copy of a recessive tumor-suppressor gene be at higher risk of developing cance...
Given that TP53 is a recessive gene and is not located on the X chromosome, why would people who inherit just one mutant copy of a recessive tumor-suppressor gene be at higher risk of developing cancer than those without the recessive gene? Given that is a recessive gene and is not located on the X chromosome, why would people who inherit just one mutant copy of a recessive tumor-suppressor gene be at higher risk of developing cancer than those without...
Please describe one well-studied example of a tumor suppressor gene
Please describe one well-studied example of a tumor suppressor gene, and highlight its normal function and how it might play a role in cancer.
 Cancer and Gene Regulation Why is a cell cycle control system needed for cell division? What happens when cells do NOT respond to the cell cycle control system and divide excessively? Tumor Proto-oncogeno (for protein that stimulates coll division) 6 Y DNA Benign Tumor= Mutation withln a control region of DNA Malignant Tumor Mutated promoter Metastasis Normal growth-stimulating protein in excess Oncogene Tumor-Suppressor Genes Proto-oncogene utled tara gese Samor-auppresr gane Many proto-oncogenes code for growth factors /Deletive nonimenig Normel grewt...
Cancer and Gene Regulation Why is a cell cycle control system needed for cell division? What happens when cells do NOT respond to the cell cycle control system and divide excessively? Tumor Proto-oncogeno (for protein that stimulates coll division) 6 Y DNA Benign Tumor= Mutation withln a control region of DNA Malignant Tumor Mutated promoter Metastasis Normal growth-stimulating protein in excess Oncogene Tumor-Suppressor Genes Proto-oncogene utled tara gese Samor-auppresr gane Many proto-oncogenes code for growth factors /Deletive nonimenig Normel grewt...
 please help me with this genetics question! thank you!
TS =Tumor Suppressor genes
O = Oncogenes
5A. (12pts) Cancer is a genetic disease. Some of the causative mutations are Tumor Suppressor genes, and others convert proto-oncogenes into Oncogenes. In the list of properties below, mark an X in the column for TS, O, or both. TS O both Causes of inherited elevated cancer risk. Leads to uncontrolled cell growth. Typically, spontaneous, gain-of-function mutations. Can cause increased DNA damage. Dominant in...
please help me with this genetics question! thank you!
TS =Tumor Suppressor genes
O = Oncogenes
5A. (12pts) Cancer is a genetic disease. Some of the causative mutations are Tumor Suppressor genes, and others convert proto-oncogenes into Oncogenes. In the list of properties below, mark an X in the column for TS, O, or both. TS O both Causes of inherited elevated cancer risk. Leads to uncontrolled cell growth. Typically, spontaneous, gain-of-function mutations. Can cause increased DNA damage. Dominant in...
 Consider a spontaneous mutation in the gene HER2, a tumor suppressor involved in cell cycle control Mutations happen at a rate of about 1 error per 109 bases per generation. A spontaneous deleterious mutation must occur in HER2 specifically, and not in another gene. The likelihood that a spontaneous mutation is deleterious and could lead to cancer depends on the number of mutable bases in HER2, the fraction of bases that will affect the function of HER2, and that both...
Consider a spontaneous mutation in the gene HER2, a tumor suppressor involved in cell cycle control Mutations happen at a rate of about 1 error per 109 bases per generation. A spontaneous deleterious mutation must occur in HER2 specifically, and not in another gene. The likelihood that a spontaneous mutation is deleterious and could lead to cancer depends on the number of mutable bases in HER2, the fraction of bases that will affect the function of HER2, and that both...
 Please help with answers to these questions
answer the ones that are straightforward like this one
Question 24 5 pts 1. Beta-catenin is classified as a proto-oncogene and APC is classified as a tumor suppressor gene. Both are important parts of the Wnt signaling pathway. Describe the relationship between beta-catenin and APC. Explain in detail why they are classified as a proto-oncogene or tumor suppressor gene. 2. Suppose you're working in a cancer research lab and discover a small molecule...
Please help with answers to these questions
answer the ones that are straightforward like this one
Question 24 5 pts 1. Beta-catenin is classified as a proto-oncogene and APC is classified as a tumor suppressor gene. Both are important parts of the Wnt signaling pathway. Describe the relationship between beta-catenin and APC. Explain in detail why they are classified as a proto-oncogene or tumor suppressor gene. 2. Suppose you're working in a cancer research lab and discover a small molecule...
 BRCAll is a tumor suppressor gene located on chromosome pair 13. Inherited mutations in BRCAII can predispose an individual to breast cancer. If Scarlet carries an inherited mutation for BRCAII and her husband Scott does not, what is the probability that their son inherited the mutated allele? O 0% o 25% O 50% O 75% O 100% Question 2 1 pts A plasmid SELECT ALL THAT APPLY. is a small circular piece of DNA independent of the chromosome is a...
BRCAll is a tumor suppressor gene located on chromosome pair 13. Inherited mutations in BRCAII can predispose an individual to breast cancer. If Scarlet carries an inherited mutation for BRCAII and her husband Scott does not, what is the probability that their son inherited the mutated allele? O 0% o 25% O 50% O 75% O 100% Question 2 1 pts A plasmid SELECT ALL THAT APPLY. is a small circular piece of DNA independent of the chromosome is a...
Most questions answered within 3 hours.
-
Investor company owns 35% of investee company voting stock and
accounts for the investment under the...
asked 5 minutes ago -
The number of major faults on a randomly chosen 1 km stretch of
highway has a...
asked 30 minutes ago -
Consider the competitive environment of Starbuck's, Progressive
Insurance, a manufacturing firm with low turnover, or a...
asked 1 hour ago -
3. Gains from trade
Consider two neighbouring island countries called Euphoria and
Contente. They each have...
asked 3 hours ago -
A business executive has the option to invest money in two
plans: Plan A guarantees that...
asked 5 hours ago -
Hello, can someone please help me answer this question?
How much heat is absorbed by a...
asked 5 hours ago -
. A marketing researcher conducted a survey of 25 shoppers
randomly selected at the local mall...
asked 5 hours ago -
Create an comprehensive response to the
following:
Antimicrobial agents work on a multitude of microbes (bacteria,...
asked 5 hours ago -
6.13 LAB: Step counter. Section 6.3.
A pedometer treats walking 2,000 steps as walking 1 mile....
asked 5 hours ago -
(14.2) A block of mass m = 10 kg riding on a frictionless
horizontal plane is...
asked 5 hours ago -
Use any search engine to search for articles about Starbucks
partnership with Tata Companies in India...
asked 5 hours ago -
Let’s say that for some reason Bank Excess Reserves suddenly
increase sharply. What effect would this...
asked 5 hours ago