
Homework Answers
The Neuromuscular Junction
The process of muscle contraction begins at the site where a motor neuron’s terminal meets the muscle fiber—called the neuromuscular junction (NMJ). Every skeletal muscle fiber in every skeletal muscle is innervated by a motor neuron at a NMJ. Excitation signals from the motor neuron are the only way to functionally activate muscle fibers to contract.
Excitation-Contraction Coupling
All living cells have membrane potentials, or electrical gradients across their membranes based on the distribution of positively and negatively charged ions. The inside of the membrane is usually around -60 to -90 mV, relative to the outside. Neurons and muscle cells can use their membrane potentials to generate and conduct electrical signals by controlling the movement of charged ions across their membranes to create electrical currents. This movement is controlled by selective opening and closing of specialized proteins in the membrane called ion channels. Although the currents generated by ions moving through these channel proteins are very small, they form the basis of both neural signaling and muscle contraction.
Both neurons and skeletal muscle cells are electrically excitable, meaning that they are able to generate action potentials. An action potential is a special type of electrical signal that can travel along a cell membrane as a wave. This allows a signal to be transmitted quickly and faithfully over long distances.
In skeletal muscle, the release of calcium to begin allowing cross-bridge formation and contraction is coupled to excitation signaling of action potentials from a motor neuron. Thus, the excitation-contraction coupling process begins with signaling from the nervous system at the neuromuscular junction (Figure 10.3.1) and ends with calcium release for muscle contraction.

(Fig10.3.1 – Motor End-Plate and Innervation: At the NMJ, the axon terminal releases ACh. The motor end-plate is the location of the ACh-receptors in the muscle fiber sarcolemma. When ACh molecules are released, they diffuse across a minute space called the synaptic cleft and bind to the receptors.)
The motor neurons that tell the skeletal muscle fibers to contract originate in the spinal cord, with a smaller number located in the brainstem for activation of skeletal muscles of the face, head, and neck. These neurons have long processes, called axons, which are specialized to transmit action potentials long distances— in this case, all the way from the spinal cord to the muscle itself (which may be up to three feet away). The axons of multiple neurons bundle together to form nerves, like wires bundled together in a cable.
Signaling begins when a neuronal action potential travels along the axon of a motor neuron, and then along the individual branches to terminate at the NMJ. At the NMJ, the axon terminal releases a chemical messenger, or neurotransmitter, called acetylcholine (ACh). The ACh molecules diffuse across a minute space called the synaptic cleft and bind to ACh receptors located within the motor end-plate of the sarcolemma on the other side of the synapse. Once ACh binds, a channel in the ACh receptor opens and positively charged ions can pass through into the muscle fiber, causing it to depolarize, meaning that the membrane potential of the muscle fiber becomes less negative (closer to zero.)
As the muscle membrane depolarizes, another set of ion channels called voltage-gated sodium channels are triggered to open. Sodium ions enter the muscle fiber, and an action potential rapidly spreads (or “fires”) along the entire membrane to initiate excitation-contraction coupling.
Things happen very quickly in the world of excitable membranes.Immeditely following depolarization of the membrane, it repolarizes, re-establishing the negative membrane potential. Meanwhile, the ACh in the synaptic cleft is degraded by the enzyme acetylcholinesterase (AChE) so that the ACh cannot rebind to a receptor and reopen its channel, which would cause unwanted extended muscle excitation and contraction.
Propagation of an action potential along the sarcolemma is the excitation portion of excitation-contraction coupling and must be coupled to the release of calcium ions for contraction. High concentrations of calcium in skeletal muscle are stored in a specialized type of smooth endoplasmic reticulum organelle called the sarcoplasmic reticulum (SR). The SR structure surrounds the myofibrils, allowing storage and release of calcium directly at sites of actin and myosin overlap. The excitation of the muscle membrane is coupled to the SR release of calcium through invaginations in the sarcolemma called T-Tubules (“T” stands for “transverse”). Because the diameter of a muscle fiber can be up to 100 μm, the T-tubules ensure that the action potential on the membrane can get to the interior of the cell and close to the SR throughout the sarcoplasm. The arrangement of a T-tubule with the membranes of SR on either side is called a triad (Figure 10.3.2).
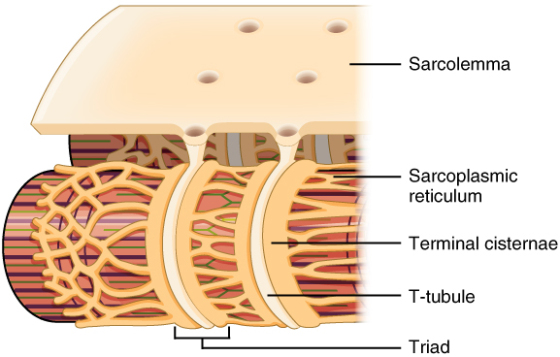
(Fig10.3.2 – The T-tubule: Narrow T-tubules permit the conduction of electrical impulses. The SR functions to regulate intracellular levels of calcium. Two terminal cisternae (where enlarged SR connects to the T-tubule) and one T-tubule comprise a triad—a “threesome” of membranes, with those of SR on two sides and the T-tubule sandwiched between them.)
Voltage-sensitive dihydropyridine receptors (DHPR) on the sarcolemma are mechanically linked to calcium channels in the adjacent SR membrane called ryanadine receptors (RyR). Through the DHPR, the action potential in the sarcolemma triggers the opening of RyR, allowing Ca++ to diffuse out of the SR and into the sarcoplasm. It is the arrival of Ca++ in the sarcoplasm that initiates contraction and shortening of sarcomeres.
Contraction and Relaxation
The sequence of events that result in the contraction of an individual muscle fiber begins with a signal—the neurotransmitter, ACh—from the motor neuron innervating that fiber. The local membrane of the fiber will depolarize as positively charged sodium ions (Na+) enter, triggering an action potential that spreads to the rest of the membrane will depolarize, including the T-tubules. This triggers the release of calcium ions (Ca++) from storage in the sarcoplasmic reticulum (SR). The Ca++ then initiates contraction, which is sustained by ATP (Figure 10.3.3). As long as Ca++ ions remain in the sarcoplasm to bind to troponin, which keeps the actin-binding sites “unshielded,” and as long as ATP is available to drive the cross-bridge cycling and the pulling of actin strands by myosin, the muscle fiber will continue to shorten to an anatomical limit.

Figure 10.3.3 – Contraction of a Muscle Fiber: A cross-bridge forms between actin and the myosin heads triggering contraction. As long as Ca++ ions remain in the sarcoplasm to bind to troponin, and as long as ATP is available, the muscle fiber will continue to shorten. Relaxation of a Muscle Fiber: Ca++ ions are pumped back into the SR, which causes the tropomyosin to reshield the binding sites on the actin strands. A muscle may also stop contracting when it runs out of ATP and becomes fatigued.
Muscle contraction usually stops when signaling from the motor neuron ends, which repolarizes the sarcolemma and T-tubules, and closes the calcium channels in the SR. Ca++ ions are then pumped back into the SR, which causes the tropomyosin to re-cover the binding sites on actin (Figure 10.3.2).
Cross-Bridge Cycling
During contraction the myosin heads of the thick filament bind to actin and pull the thin filament which shortens the sarcomere and produces force. However, the length of the mysoin hinge region allows each myosin head to only pull a very short distance before it must reset to pull again. For thin filaments to continue to slide past thick filaments during muscle contraction, myosin heads must pull the actin at the binding sites, detach, re-cock, attach to more binding sites, pull, detach, re-cock, etc. This repeated movement is known cross-bridge cycling and is dependent on ATP (Figure 10.3.4). Restoring the myosin head to position to pull on actin requires energy which is provided by ATP.

(Fig10.3.4 – Skeletal Muscle Contraction: (a) The active site on actin is exposed as calcium binds to troponin. (b) The myosin head is attracted to actin, and myosin binds actin at its actin-binding site, forming the cross-bridge. (c) During the power stroke, the phosphate generated in the previous contraction cycle is released. This results in the myosin head pivoting toward the center of the sarcomere, after which the attached ADP and phosphate group are released. (d) A new molecule of ATP attaches to the myosin head, causing the cross-bridge to detach. (e) The myosin head hydrolyzes ATP to ADP and phosphate, which returns the myosin to the cocked position.)
Each myosin head has a region that binds to actin and a region that binds to ATP. Myosin cannot release from actin until ATP also binds, and the hydrolysis of ATP into adenosine diphosphate (ADP) and inorganic phosphate (Pi) then releases energy needed for the myosin head to reposition or re-cock.
Cross-bridge formation occurs when the myosin head attaches to the actin while adenosine diphosphate (ADP) and inorganic phosphate are still bound to myosin (Figure 10.3.4a,b). Pi is then released, causing myosin to form a stronger attachment to the actin, after which the myosin head moves toward the M-line, pulling the actin along with it. As actin is pulled, the filaments move approximately 10 nm toward the M-line. This movement is called the power stroke, as movement of the thin filament occurs at this step (Figure 10.3.4c). In the absence of ATP, the myosin head will not detach from actin.
ATP binding causes the myosin head to detach from the actin (Figure 10.3.4d). After this occurs, ATP is converted to ADP and Pi by the intrinsic ATPase activity of myosin. The energy released during ATP hydrolysis changes the angle of the myosin head into a cocked position (Figure 10.3.4e). The myosin head is now in position for further movement.
When the myosin head is cocked, myosin is in a high-energy configuration. This energy is expended as the myosin head moves through the power stroke, and at the end of the power stroke, the myosin head is in a low-energy position. After the power stroke, ADP is released; however, the formed cross-bridge is still in place, and actin and myosin are bound together. As long as ATP is available, it readily attaches to myosin, the cross-bridge cycle can recur, and muscle contraction can continue.
Each thick filament of roughly 300 myosin molecules has multiple myosin heads, and many cross-bridges form and break continuously during muscle contraction. Multiply this by all of the sarcomeres in one myofibril, all the myofibrils in one muscle fiber, and all of the muscle fibers in one skeletal muscle, and you can understand why so much energy (ATP) is needed to keep skeletal muscles working. In fact, it is the loss of ATP that results in the rigor mortis observed soon after someone dies. With no further ATP production possible, there is no ATP available for myosin heads to detach from the actin-binding sites, so the cross-bridges stay in place, causing the rigidity in the skeletal muscles.
Sources of ATP
ATP supplies the energy for muscle contraction to take place. In addition to its direct role in the cross-bridge cycle, ATP also provides the energy for the active-transport Ca++ pumps in the SR. Muscle contraction does not occur without sufficient amounts of ATP. The amount of ATP stored in muscle is very low, only sufficient to power a few seconds worth of contractions. As it is broken down, ATP must therefore be regenerated and replaced quickly to allow for sustained contraction. There are three mechanisms by which ATP can be regenerated: creatine phosphate metabolism, anaerobic glycolysis, fermentation and aerobic respiration.
Action of Sarin and BOTOX
Like other nerve agents, sarin targets an enzyme within the body's neuromuscular junctions, where nerves meet muscles. Usually, this enzyme deactivates the nerve-signaling molecule acetylcholine. This can lead acetylcholine to build up in the muscles, cause excessive twitching and then result in paralysis.Intramuscular administration of botulinum toxin acts at the neuromuscular junction to cause muscle paralysis by inhibiting the release of acetylcholine from presynaptic motor neurons.
Add Answer to:
Short answer (answer on the back of page or attach pages): List the steps in order...
Put these steps in neuromuscular junction synaptic transmission and excitation-contraction coupling in chronological order. A. Ryanodine...
Put these steps in neuromuscular junction synaptic transmission and excitation-contraction coupling in chronological order. A. Ryanodine receptors open B. Voltage-gated Na+ channels open in the muscle cell membrane near the neuromuscular junction C. Tropomyosin rotates into the groove of the thin filament (out of the way of the myosin binding site) D. Depolarization of motoneuron axon terminal E. Ca2+ dissociates from troponin F. [Ca2+] rises in the muscle cell cytoplasm G. An EPSP occurs in the muscle cell H. Myosin...
1. A. Name the three planes and the positions they each describe to identify a unique...
1. A. Name the three planes and the positions they each describe to identify a unique position in the human body. B. Name the two major ventral body cavities plus the major organs found in them. C. Name the cavities that the heart and lungs reside in. D. Finally, list the six levels of organization in nature. 2. A. Describe the three components of an atom in terms of charge and location. Define atomic mass and atomic number. B. For...
1. According to the paper, what does lactate dehydrogenase (LDH) do and what does it allow...
 1. According to the paper, what does lactate dehydrogenase
(LDH) do and what does it allow to happen within the myofiber? (5
points)
2. According to the paper, what is the major disadvantage of
relying on glycolysis during high-intensity exercise? (5
points)
3. Using Figure 1 in the paper, briefly describe the different
sources of ATP production at 50% versus 90% AND explain whether you
believe this depiction of ATP production applies to a Type IIX
myofiber in a human....
1. According to the paper, what does lactate dehydrogenase
(LDH) do and what does it allow to happen within the myofiber? (5
points)
2. According to the paper, what is the major disadvantage of
relying on glycolysis during high-intensity exercise? (5
points)
3. Using Figure 1 in the paper, briefly describe the different
sources of ATP production at 50% versus 90% AND explain whether you
believe this depiction of ATP production applies to a Type IIX
myofiber in a human....
 1. According to the paper, what does lactate dehydrogenase
(LDH) do and what does it allow to happen within the myofiber? (5
points)
2. According to the paper, what is the major disadvantage of
relying on glycolysis during high-intensity exercise? (5
points)
3. Using Figure 1 in the paper, briefly describe the different
sources of ATP production at 50% versus 90% AND explain whether you
believe this depiction of ATP production applies to a Type IIX
myofiber in a human....
1. According to the paper, what does lactate dehydrogenase
(LDH) do and what does it allow to happen within the myofiber? (5
points)
2. According to the paper, what is the major disadvantage of
relying on glycolysis during high-intensity exercise? (5
points)
3. Using Figure 1 in the paper, briefly describe the different
sources of ATP production at 50% versus 90% AND explain whether you
believe this depiction of ATP production applies to a Type IIX
myofiber in a human....
Most questions answered within 3 hours.
-
Potassium permanganate(KMNO4)is has a solubility of 6.4 g/ 100 g
of water at 20ºC, and 250...
asked 43 seconds ago -
Given the following information:
acetic acid
CH3COOH
Ka = 1.8×10-5
triethylamine
(C2H5)3N
Kb = 5.2×10-4
(1)...
asked 3 minutes ago -
51.
As the marginal propensity to expend rises, the multiplier:
decreases.
is impossible to determine.
increases....
asked 7 minutes ago -
The Baldwin Company currently has the following balances on their
balance sheet:
Total
Liabilities
$69,309
Common...
asked 10 minutes ago -
A quiet town in Kansas has 10 people, all of whom have the same
preferences. There...
asked 15 minutes ago -
Summarize what an organization needs from a leader.
a. Analyze what might happen to an organization...
asked 18 minutes ago -
How would one critically evaluate an organizations marketing
strategies from the viewpoint of its consumers, as...
asked 17 minutes ago -
Given a standardized normal distribution (with μ = 0 and a σ =
1), what is...
asked 17 minutes ago -
Company XYZ know that replacement times for the quartz time
pieces it produces are normally distributed...
asked 32 minutes ago -
A) Write the acid-base reaction that occurs between Na2HPO4 and
NaOH
B) Write the possible reactions...
asked 46 minutes ago -
What advantages does the Natural Law Theory have in comparison
with the Divine Command Theory? Explain...
asked 52 minutes ago -
A diver comes off a board with arms straight up and legs
straight down, giving her...
asked 51 minutes ago